About Me
Hello there Earthlings! My name is Jacob Schimp and I'm a rising senior at the University of Illinois at Urbana-Champaign
majoring in Physics. This summer I had the opportunity to work with Dr. Alvin Bayliss on
nonlinear crowding effects in cyclical ecosystems.
Contact info: jschimp2@illinois.edu
Background
In population dynamics, the crowding effect, representing intra-species competition, has long been recognized as a pivotal determinant of population growth. Traditionally, this effect is assumed to follow a linear relationship concerning the population’s growth, wherein an increase in population density leads to a proportional increase in the crowding contribution to the growth rate. However, ecological systems often exhibit complexities that demand a more nuanced understanding of competitive interactions via crowding, Gavina et al. (2018). In the context of two-species competition, the Lotka-Volterra model has extensively informed our understanding of classical systems, Grover (1997). Assuming an appropriate non-dimensionalization, the equations of the Lotka-Volterra equations are:
$\displaystyle{\frac{du}{dt} = u(1- ${\frac{u}{1 \pm B}}$$-\alpha v),}
$\displaystyle{\frac{dv}{dt} = rv(1-${\frac{v}{1 \pm B}}$$ - \beta u) \, ,}$
where the terms in red represent intra-species competition (crowding). Here 𝑟 represents the ratio of the natural birthrate of species 𝑣 to that of species 𝑢, and 𝛼 and 𝛽 characterize the inter-species competition. When 𝐵 = 0 (linear crowding growth) this model has long provided insight and understanding of population dynamics in two-species ecosystems, Wang et al. (2022), such as determining conditions when both species can coexist (biodiversity) or only one of the species can survive (bioselectivity). However, as we extend our inquiry to nonlinear crowding, we modify the crowding terms (terms in red) in (1) and (2) ($B$ > 0). We say that a community crowds faster than linear, $F$, when we take $-$ in the denominator of the crowding term and a community crowds slower than linear, $S$, when we take $+$ in the denominator.
For the classical model (1) and (2), and taking α = β for simplicity, there are four critical points:1. b00 = (0, 0) (both species goes extinct)
2. b10 = (1, 0) (species u wins and v loses)
3. b01 = (0, 1) (species v wins and u loses)
4. buv = (u∗, v∗) (species u and v coexist)
We solved and reviewed the stability of the system by checking if the eigenvalues of the associated Jacobian matrix were positive or negative for each critical point. Coexistence will be stable if α < 1 while the single species states will be stable for α > 1 (bistability regime). The extinction state b00 is always unstable.
FINDINGS
Symmetric Case of Two Species
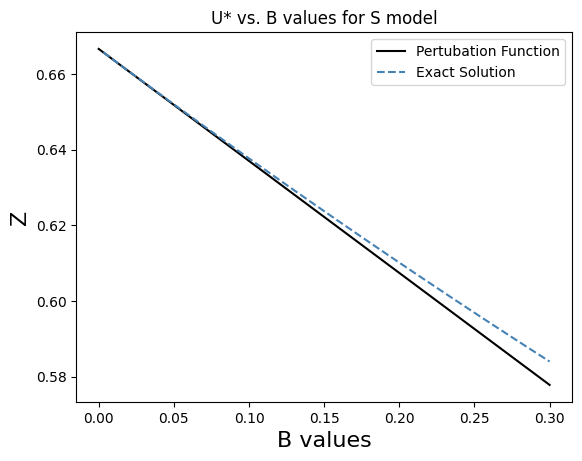
We include nonlinear crowding, with parameter, B, where 𝐵 > 0 and, for simplicity, we take 𝑟 = 1. For F and S communities, coexistence will be stable under the exact reverse of the stability conditions for the single- species case, i,e., when 𝛼 < 1 + 𝐵 for F , and 𝛼 < 1 − 𝐵 for 𝑆. When these conditions are satisfied both species reach a stable state 𝑢∗ over time, independent of initial conditions. The transition point where species shift from single-species dominance to coexistence is α = 1 + B for 𝐹 communities and 𝛼 = 1 − 𝐵 for 𝑆 communities. Our model of nonlinear crowding growth is controlled by 𝐵. When 𝐵 > 0 the terms in red now describe our model of nonlinear crowding. The crowding term can incorporate faster than linear growth (− sign in the red denominator - F community) or slower than linear growth (+ sign in the red denominator - S community). For simplicity we describe our results only for the symmetric case when β = α . When 𝐵 = 0 the classical result is that coexistence will be stable if 𝛼 < 1 while the single species states will be stable for 𝛼 > 1 (bistable regime). For 𝑆 communities the single-species states are stable if 𝛼 > 1 − 𝐵 while for 𝐹 communities the(window for coexistence, e.g., biodiversity, shrinks) single species states are stable if 𝛼 > 1 + 𝐵(window for coexistence, e.g., biodiversity). In the symmetrical case the coexistence point (𝑢∗, 𝑣∗) satisfies 𝑣∗ = 𝑢∗ where 𝑢∗ is a root of a quadratic equation. The two roots are:
$\displaystyle{u_{*} = -\frac{(1+\alpha-B) \pm \sqrt{(1+\alpha-B)^2+4(\alpha B)}}{2 \alpha B}}$We have developed perturbation and asymptotic expansions for the roots for small 𝐵.
Asymmetric Case of Two Species
For the asymmetric case, we take 𝛼 ≠ 𝛽 and the equations are in the form outlined in the symmetric case. For 𝐹 communities, we showed that for the 𝑏10 (𝑢 wins, 𝑣 loses) state to be stable, the parameters must satisfy 𝛽 > 1 + 𝐵. Similarly, for the 𝑏01 (𝑣 wins, 𝑢 loses) state to be stable, the parameters must satisfy 𝛼 > 1 + 𝐵. Similarly, for 𝑆 communities, the transition point for the 𝑏01 state is 𝛽 = 1 + 𝐵 and for the 𝑏10 state 𝛽 = 1 − 𝐵 . The coexistence state for the asymmetric case (𝑢∗, 𝑣∗) satisfies the coupled quadratic equations which cannot be solved explicitly. As an example, for 𝐹 communities the system is:
$\displaystyle{0=1- (\beta+1)u_{*}-\alpha v_{*} +B \alpha u_{*}v_{*}}$
$\displaystyle{0=1-(\beta+1)v_{*}-\beta u_{*} +B\beta u_{*}v_{*}}$We have demonstrated coexistence 𝐵 is small, but due to the complicated nature of the coupled equations, we have not been able to analytically determine transition values when crowding growth is nonlinear.

Symmetric Case of Three Species
We next studied a three-species cyclic system. Such systems are often referred to as rock-paper-scissors (𝑅𝑃𝑆) system because of the clear analogy with the classical children’s game, Bayliss et al. (2019). After appropriate non- dimensionalization and assuming that all natural birthrates are equal, the system of ODEs governing this ecological system takes the following form:
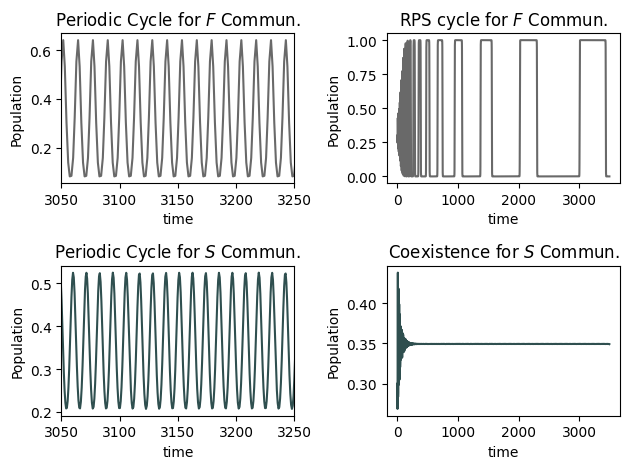
We next considered the impact of the nonlinear crowding effect ($B>0$) and considered both faster and slower than linear growth communities ($F$ and $S$ communities, respectively). We took $B = 0.1\,$. For specific ranges of $\alpha$, the interval $[2.1,2.2]$ for $F$ communities and [1.60,1.8] for S communities), a new behavior emerges - a window of periodic solutions, describing a periodic change in dominance for the system. In this scenario, each species dominates the ecosystem for a specific duration, and as time approaches infinity, the time intervals for each species’ dominance remain constant. This periodic window is not observed for linear crowding ($B=0$).
Conclusion
In summary, the findings of this research shows the dynamics of nonlinear crowding in shaping the stability of coexistence within ecosystems for $N= 2,3$ species. We found that for faster-than-linear crowding ($F$ community), parameters windows contract, emphasizing the constraints imposed on coexistence. Conversely, in scenarios of slower-than-linear crowding, the parameter windows expand, offering greater possibilities for stable coexistence. Moreover, for the case of two-species systems ($N=2$), we derived a perturbation expansion for the coexistence state in symmetric competitive communities. For three-species systems ($N=3$), our study revealed a deviation from the $RPS$ cycle which was interrupted by a periodic behavior at a range of $\alpha$ values for both $F$ and $S$ communities. In summary, our research offers significant insights into ecological interactions and dynamics that contribute to a deeper understanding of different ecosystems that may exhibit nonlinear crowding effects.
Acknowledgements
We recognize the Potawatomi, Ojibwe, and Odawa tribes as the traditional stewards of this land. We are committed to learning about and respecting the deep history and contributions of these tribes and nations. We offer our respect to the elders, past and present, and to all Indigenous peoples present today. This material is based upon work supported by the National Science Foundation under grant No. AST 2149425, a Research Experience for Undergraduates (REU) grant awarded to CIERA at Northwestern University. Any opinions, findings, conclusions, or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.